健康な体づくりを。
一緒に解決します。

CONCEPT
腸元氣堂は、生活習慣の乱れを整える腸もみサロンです
腸の不調でもある便秘と言っても、下剤の使い過ぎ、食生活の乱れ、ストレス、腸内細菌のバランスの乱れなど、原因は様々です。
医療機関ではそれらを考慮することなく、下剤を処方することで対応していますが、
本当に重要なのは「なぜそのような状況になっているか」原因を探り、それを取り除いてあげること。
腸元氣堂では、便秘の改善はもとより、なぜそのような状況になったのか、
これからどういう風にしていけばいいのか、一人ひとりのご相談にきちんと耳を傾け、一緒に悩み、考えていきます。
銀座、綾瀬、中延など東京都内とつくばに腸もみ店舗を展開しております。
最近、腸の大切さが認知され、それと併せて腸もみの認知度もアップしてきました。
地方から東京に用事で来られるついでに腸もみを受けたいと腸元氣堂にご来店頂くことも増えてきました。
東京3店舗、銀座、綾瀬、中延と行きやすい場所にあるのも理由のひとつかと思います。
腸元氣堂で腸もみを受けるメリットとして、東京に住んでいなくても、
普段の食事や運動などに関して湧き上がってきた疑問や質問をLINEやショートメールなどで その人に合ったやりやすい方法でご提案する時もあります。
便秘解消やむくみ、ぽっこりお腹解消はこうした日常で起こる疑問を解決して、
自律神経を整えることが大事だと思います。
続けるには無理なく楽しく実践できることが大事だと思うので、
その点も気をつけていることのひとつです。
腸もみってなに?
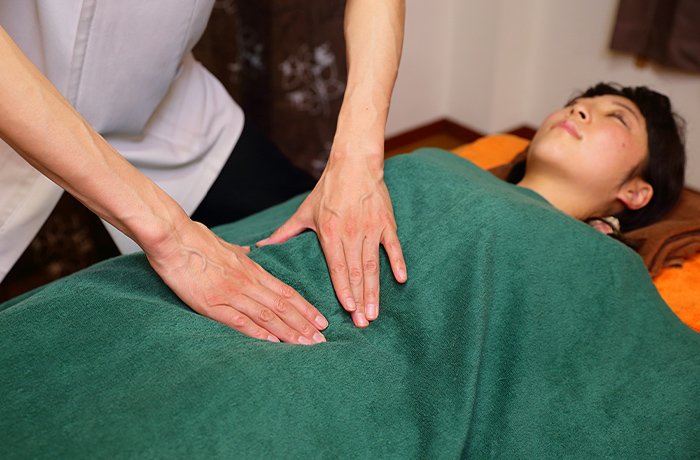
腸もみとは硬くなった腸や動きが鈍くなってしまった腸を刺激することで、その機能を回復させる治療法のこと。たとえば、女性を中心に多くの方がお悩みの「便秘」に対して効果的です。便秘に明確な定義はありませんが一般的には「3日便通がない状態」をさすと言われています。非常に苦しい状態ですが、医療機関を受診しても「下剤を処方して終わり」という一時しのぎのやり方ばかり。根本的な解決には至らない場合がほとんどです。
腸もみの効果は?
便秘が肩こり、腰痛などの、他の体の不調の原因となっている場合も少なくありません。それは便秘になることで腸が癒着したり、炎症を起こした結果、筋肉を不自然に引っ張ったり、他の臓器を圧迫するためです。腸もみをすることで、これらの改善も期待できます。
また、先に挙げた通り、血液がキレイになることで病気の予防につながったり、セロトニンなどのホルモンが分泌されることで安眠につながったり、善玉菌が増えることで代謝が高まったり、心身に良い影響が期待できます。実際、患者様の中には便秘の改善以外にも「花粉症が改善された」「ぽっこりお腹がへこんだ」「体重が落ちた」など嬉しいお声を多数頂いています。
腸もみサロン腸元氣堂って
腸元氣堂は、生活習慣の乱れを整える腸もみサロンです
健康における腸の環境はとても大切です。
腸元氣堂は便秘の改善はもとより、なぜそのような状況になったのか、これからの対策はどうすれば良いのかなど一人ひとりのご相談に真摯に耳を傾け、一緒に悩み、考えていきます。
日常で起こる疑問を解決して、便秘解消やむくみ、ぽっこりお腹など自律神経を整えて健康的な日常を提供して行ければ幸いであると考えております。
東京都内の銀座、綾瀬、中延やつくばに店舗を展開しております。東京に住んでいなくても、普段の食事や運動などに関して湧き上がってきた疑問や質問をLINEやショートメールなどで その人に合ったやりやすい方法でご提案する時もあります。
お悩みがある際は、お問い合わせよりお気軽にご相談ください。

腸もみマスターが
サポートします!

腸もみマスター
江川 泰志
保有資格
インテスタージュ®トリートメント腸セラピー
免疫美容
栄養医学指導師
悩みに寄り添い、
最善を尽くす。
様々な症状でお困りの方の手助けをしたい。
私と腸もみとの出会いは自身の便秘解消。私の場合、食べ過ぎが原因でしたが、便秘解消以上に身体が軽くなる実感があり、多くの方に伝えていきたいと思ったことが、キッカケです。
いろいろと知る中で、世の中の常識が実は間違っていて、そのために要らない苦労をしている人がたくさんいることにも知りました。
腸元氣堂は、症状や重症度問わず、患者様個々のお悩みに寄り添い、「何とかしてあげたい」という思いのもと、できる最善を尽くすことを心掛けています。






 腸もみを体験したお客様のお喜びの声
腸もみを体験したお客様のお喜びの声
長年悩んでいた頭痛やむくみ・冷えの原因が、まさか腸だったなんて!




大学生の頃からすぐにお腹が痛くなり、ベッドから出られないという日がよくありました。とくにストレスが溜まっているとかそういう感じもなく原因がわかりませんでした。 ですが、施術をしていただくと腸が硬くなっていると言われてビックリしました。腸って硬くなるんだ!と思い驚きましたが、それより施術の効果を実感してまたビックリ! 胃が痛くなる以外に冷え性に悩んでいましたが、それも今では改善されてきています。もうお腹が痛くて動けない・・・という思いはしたくないので、今後も院長にお世話になりたいと思います!




お知らせ
- 2023年12月27日 年末年始の営業お知らせ
- 2023年9月13日 【メディア掲載情報】おとなの週末10月号に当店が掲載されました
- 2023年8月10日 お盆も営業します。
- 2023年8月10日 【銀座店】店舗移転のお知らせ ※8/17(木)〜
- 2023年4月26日 ゴールデンウイーク中も営業します。
新型コロナウイルス感染防止対策について
当店での新型コロナウイルス対策は下記の対策を行います。
◆施術者のマスク着用、施術前後の手の消毒
◆お客様の触れる可能性がある所
(ドアノブ・スリッパ・お着替えをお入れしている籠・ハンガー・施術ベッド・ソファー・クッション・お手洗い周り・床等)の消毒
◆施術中も窓(2〜3箇所)を開けての換気
◆プラズマクラスター空気清浄機使用
(プラズマクラスターとは、菌やウイルスに作用して風邪やインフルエンザ予防に効果が期待できると言われています)
◆殺菌作用があるアロマ使用
今後とも安心してお客様にご利用いただけますよう努めてまいります。


生理不順・便秘・むくみ・冷え・ストレス・不眠・肩こり・腰痛・ダイエットなど、体の不調でお悩みなら、まずはお電話ください!
初回20%OFF
総合受付 9:00~23:00(最終受付22:00)
※店舗によって営業時間が異なります